








В генетической инженерии для конструирования рекомбинантных ДНК используется множество различных ферментов. В клетке межнуклеотидные связи в ДНК и РНК расщепляются нуклеазами. В генетической инженерии используют эндонуклеазы рестрикции — рестриктазы. Эти ферменты распознают в молекулах ДНК определенные нуклеотидные последовательности из четырех, пяти или шести остатков и поэтому расщепляют эту кислоту на сравнительно небольшое число строго определенных фрагментов.
Еще в 1953 г. было обнаружено, что ДНК определенного штамма Е. coliу введенная в клетки другого штамма (например, ДНК штамма В в клетки штамма С), не проявляет, как правило, генетической активности, так как быстро расщепляется на мелкие фрагменты. В 1966 г. установлено, что это явление связано со специфической модификацией хозяйской ДНК. В 1969 г. были открыты:
- специфический модифицирующий фермент,
- метилирующий ДНК,
- рестриктаза, которая расщепляла неметилированную ДНК.
Однако этот фермент не был высокоспецифичен по отношению к определенной последовательности этой кислоты. Вскоре была выделена первая рестриктаза, которая расщепляла строго определенную последовательность ДНК.
Рестриктазы принято именовать по названию бактерий, из которых их выделяют. Так, название EcoRl свидетельствует о том, что этот фермент получен из Escherichiacoli. Каждый фермент узнает определенную 4 — 7-членную последовательность в двухцепочечной ДНК. Разрезание последней по этим сайтам приводит к образованию:
- либо тупых (например, при действии рестриктазы Hpal),
- либо липких, т. е. перекрывающихся (например, BamHI), концов.
Для конструирования гибридных молекул особенно удобны последние.
Если допустить, что нуклеотиды распределены по молекуле ДНК совершенно случайным образом, можно рассчитать частоту встречаемости участка узнавания для данной рестриктазы. В каждой нуклеотидной позиции молекулы ДНК с одинаковой вероятностью может оказаться один из 4 нуклеотидов (A, G, С, Т), поэтому фермент, узнающий четырехзвенную последовательность, будет находить одну мишень из 256 пар оснований. В то же время фермент, различающий шестизвенную последовательность, определит специфический участок узнавания 1 раз из 4 096 пар оснований.
Любой фрагмент ДНК обладает характерным расположением сайтов узнавания различных рестриктаз, что позволяет строить так называемые рестриктазные карты. Небольшие геномы, например, геномы вирусов, митохондрий, хлоропластов, или части крупных геномов могут быть таким образом расщеплены на определенное число рестрикционных фрагментов. В дальнейшем эти фрагменты можно выделить, чтобы определить, каким генам и (или) регуляторным участкам они соответствуют. Их также можно использовать, чтобы синтезировать гетерологичные рекомбинантные молекулы, встроив, например, какой-либо фрагмент в плазмиду и размножив его с помощью Е. coli. Получение коротких фрагментов ДНК необходимо и для установления последовательности оснований в более крупных фрагментах.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Рестриктазы бывают:
- мелкощепящими,
- крупнощепящими.
Первые узнают тетрануклеотид и вносят в молекулы гораздо больше разрывов, чем вторые, распознающие последовательность из шести нуклеотидных пар.
Для создания рекомбинантных ДНК используют также РНК-зависимые ДНК-полимеразы (обратные транскриптазы, ревертазы), синтезирующие ДНК на матрице мРНК. Применяются и ферменты, соединяющие концы фрагментов ДНК, т. е. ДНК-лигазы, выделяемые из Е. coliи фага Т4. Для изменения структуры концов фрагментов ДНК используют:
- нуклеазу BamI,
- экзонуклеазу III из E.coli,
- нуклеазу SIиз Aspergillusorysae.
Субстратом для таких ферментов являются:
- однотяжевые РНК и ДНК,
- двутяжевые ДНК, которые распадаются до 5'-фосфорилированных олигонуклеотидов и нуклеозид-5'-фосфатов.
Для приготовления гибридизационных проб используется ДНК- полимераза I из Е. coli.
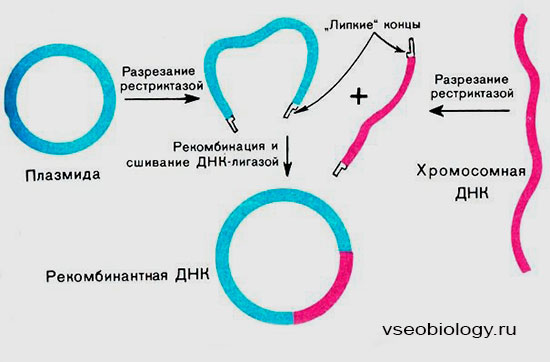
Существует три способа конструирования рекомбинантных ДНК.
- Первый — объединение ДНК-фрагментов по липким концам. Как мы уже говорили, некоторые рестриктазы способны образовывать их при специфическом разрезании ДНК. Используя это, необходимый ген выделяют из нее при помощи какой-либо рестриктазы. Ею же обрабатывают ту ДНК, к которой хотят присоединить ген. В результате как на выделенном гене, так и на другой ДНК образуются комплементарные липкие концы.
- Второй способ — коннекторный. Существуют рестриктазы, образующие при специфическом расщеплении ДНК тупые концы. Для гена, полученного таким образом, необходимо пришить липкие концы. Это достигается приливанием, например, смеси аденозина или тимина.
- Третий способ, линкерный, предполагает, что при необходимости тупые концы могут быть превращены в липкие. Для этого к первым присоединяют двухцепочечные последовательности (линкеры) с участками узнавания рестриктазы, дающей липкие концы.