








На цитоплазматических мембранах практически всех клеток макроорганизма обнаруживаются антигены гистосовместимости. Большая часть из них относится к системе главного комплекса гистосовместимости, или MHC (от англ. Main Hystocompatibility Complex). При пересадках органов возникает проблема совместимости тканей, связанная со степенью их генетического родства, реакциями отторжения чужеродных аллогенных и ксеногенных трансплантатов, т.е. проблемами трансплантационного иммунитета. Существует ряд тканевых антигенов. Трансплантационные антигены во многом определяют индивидуальную антигенную специфичность организма. Сопокупность генов, определяющих синтез трансплантационных антигенов, получила название главной системы гистосовместимости. Установлено, что антигены гистосовместимости играют ключевую роль в осуществлении специфического распознавания «свой-чужой» и индукции приобретенного иммунного ответа, определяют совместимость органов и тканей при трансплантации в пределах одного вида и другие эффекты. Большая заслуга в изучении MHC принадлежит Дж. Доссе, П. Догерти, П. Гореру, Г. Снеллу, Р. Цинкернагелю, Р.В. Петрову, ставшими основоположниками иммуногенетики.
Впервые MHC был обнаружен в 60-х годах ХХ века в опытах на генетически чистых (инбредных) линиях мышей при попытке межлинейной пересадки опухолевых тканей (П. Горер, Г. Снелл). У мышей этот комплекс получил название Н-2 и был картирован в 17-й хромосоме.
У человека MHC был описан несколько позже в работах Дж. Доссе. Его обозначили как HLA (от англ. Human Leukocyte Antigen), так как он ассоциирован с лейкоцитами. Биосинтез HLA определяется генами, локализованными сразу в нескольких локусах короткого плеча 6-й хромосомы.
MHC имеет сложную структуру и высокую полиморфность. Антигены гистосовместимости представляют собой гликопротеины, прочно связанные с цитоплазматической мембраной клеток. Их отдельные фрагменты имеют структурное сходство с молекулами иммуноглобулинов и поэтому относятся к единому суперсемейству.
Различают два основных класса молекул MHC (I и II), которые объединяют множество сходных по структуре антигенов, кодируемых множеством аллельных генов. На клетках индивидуума могут одновременно экспрессироваться не более двух разновидностей продуктов каждого гена MHC. MHC I класса индуцирует преимущественно клеточный иммунный ответ, а MHC II класса - гуморальный.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
MHC I класса состоит из двух нековалентно связанных полипептидных цепей (α и β) с разной молекулярной массой. α-Цепь имеет внеклеточный участок с доменным строением (α1-, α2- и а3-домены), трансмембранный и цитоплазматический. β-Цепь представляет собой β2-микроглобулин, адгезированный на α-домен после экспрессии α-цепи на цитоплазматической мембране клетки. α1- и α2-Домены α-цепи формируют щель Бъеркмана - участок, ответственный за сорбцию и презентацию молекул
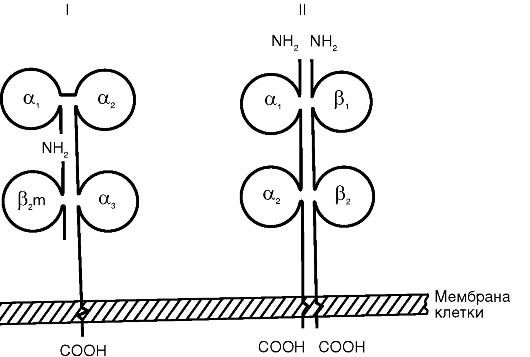
Схема строения антигенов главного комплекса гистосовместимости: I - МНС I класса; II - МНС II класса антигена.
Щель Бъеркмана MHC I класса вмещает нанопептид, который легко выявляется специфическими антителами.
Сборка комплекса MHC I класса - антиген протекает внутриклеточно непрерывно в эндоплазматическом ретикулуме. В его состав включаются любые эндогенно синтезированные пептиды, в том числе вирусные, куда они переносятся из цитоплазмы при помощи особого белка, протеосомы. Включенный в комплекс пептид придает структурную устойчивость MHC I класса. В его отсутствие функцию стабилизатора выполняет шаперон (калнексин).
MHC I класса экспрессируются на поверхности практически всех клеток, кроме эритроцитов и клеток ворсинчатого трофобласта (профилактика отторжения плода). Плотность MHC I класса достигает 7000 молекул на клетку, и они покрывают около 1% ее поверхности. Для них характерна высокая скорость биосинтеза - процесс завершается за 6 ч. Экспрессия MHC I класса усиливается под влиянием цитокинов, например, γ-интерферона.
В настоящее время у человека различают более 200 различных вариантов HLA I класса. Они кодируются генами, картированными в трех основных сублокусах 6-й хромосомы и наследуются и проявляются независимо: HLA-A, HLA-B и HLA-C.
Локус А объединяет более 60 вариантов, В - 130, а С - около 40. Независимое наследование генов сублокусов в популяции формирует бесконечное множество неповторяющихся комбинаций HLA I класса. Каждый человек строго уникален по набору антигенов гистосовместимости, исключение составляют только однояйцовые близнецы.
Основная биологическая роль HLA I класса - они определяют биологическую индивидуальность (биологический паспорт) и являются маркерами «своего» для иммунокомпетентных клеток. Заражение клетки вирусом или ее мутация изменяют структуру HLA I класса, что является сигналом для активации Т-киллеров (CD8+-лимфоциты) к уничтожению объекта.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
HLA I класса выявляют на лимфоцитах в реакции микролимфоцитолиза со специфическими сыворотками, которые получают от многорожавших женщин, пациентов после массивной гемотрансфузии, а также с использованием моноклональных антител.
В структуре и функции MHC II класса есть ряд принципиальных отличий. Комплекс образован двумя нековалентно связанными полипептидными цепями (α и β), имеющими сходное доменное строение. Обе цепи являются трансмембранными пептидами и «заякорены» в цитоплазматической мембране. Щель Бъеркмана в MHC II класса образована одновременно обеими цепями. Она вмещает олигопептид размером 12-25 аминокислотных остатков, недосягаемый специфическими антителами. MHC II класса включает в себя пептид, захваченный из внеклеточной среды путем эндоцитоза, а не синтезированный самой клеткой. Молекулы МНС II класса экспрессируются на поверхности ограниченного числа клеток: дендритных, В-лимфоцитах, Т-хелперах, активированных макрофагах, тучных, эпителиальных и эндотелиальных клетках. Обнаружение MHC II класса на нетипичных клетках расценивается в настоящее время как иммунопатология. Биосинтез MHC II класса протекает в эндоплазматическом ретикулуме и экспрессируется на цитоплазматической мембране клетки в течение 1 ч после эндоцитоза антигена. Экспрессия комплекса может быть усилена γ-интерфероном и снижена простагландином Е2.
По имеющимся данным, человеческому организму свойствен чрезвычайно высокий полиморфизм HLA II класса, который в большей степени определяется особенностями строения β-цепи. В состав комплекса входят продукты трех основных локусов: HLA- DR, DQ и DP. При этом локус DR объединяет около 300 аллельных форм, DQ - около 400, а DP - около 500.
Наличие и тип MHC II класса определяют в серологических (микролимфоцитотоксический тест) на В-лимфоцитах и клеточных реакциях иммунитета (смешанная культура лимфоцитов). Специфические антитела к MHC II класса получают так же, как и к I классу. Тестирование в смешанной культуре лимфоцитов позволяет выявить минорные компоненты MHC II класса, не определяемые серологически.
MHC II класса участвуют в индукции приобретенного иммунного ответа. Фрагменты молекулы антигена экспрессируются на цитоплазматической мембране особой группы клеток, которая получила название антигенпрезентирующих. Основными являются дендритная клетка, макрофаг и В-лимфоцит. Структура MHC II класса с включенным в него пептидом в комплексе с кофакторными молекулами CD-антигенов воспринимается и анализируется Т-хелперами (CD4+-лимфоциты). В случае распознавания чужеродности Т-хелпер начинает синтез соответствующих иммуноцитокинов, и включается механизм специфического иммунного реагирования: пролиферация и дифференцировка антигенспецифических клонов лимфоцитов.
Помимо описанных выше антигенов гистосовместимости, идентифицирован III класс молекул MHC. Локус, содержащий кодирующие их гены, вклинивается между I и II классами и разделяет их. К MHC III класса относятся некоторые компоненты комплемента (С2, С4), белки теплового шока, факторы некроза опухоли и др.