








Первичное распознавание чужеродного антигена осуществляется функционально неподготовленными наивными Т-клетками, хотя внутритимусная дифференцировка уже определила основные свойства этих клеток, что отражается в их делении на субпопуляции.
Созревание наивных Т-клеток в зрелые функционально активные (армированные) Т-клетки происходит в лимфоидной ткани. Место проникновения патогена (в более широком смысле - антигена) в организм, как правило, удалено от лимфоидных образований. С током лимфы антиген попадает в ближайшие лимфатические узлы. Если антиген оказывается в кровотоке, то он оседает в селезенке. Локализация антигена на слизистых приводит к его миграции в лимфоидную ткань, ассоциированную с этими слизистыми покровами: в пейеровы бляшки, миндалины .
В периферической лимфоидной ткани имеются три типа специализированных клеток, которые способны усваивать антиген и представлять его в иммуногенной форме на своей поверхности для распознавания Т-клетками. Это - макрофаги, дендритные клетки и В-клетки. Все они получили общее название антигенпрезентирующих клеток (АПК).
Макрофаги не имеют гистологически определенного места локализации и широко представлены по всей лимфоидной ткани. Дендритные клетки связаны с Т-зоной лимфатических узлов. В-клетки концентрируются в фолликулах.
Функция этих типов клеток - представление антигенных пептидов в комплексе с молекулами MHC класса I или II, т.е. придание проникшему антигену иммуногенных свойств.
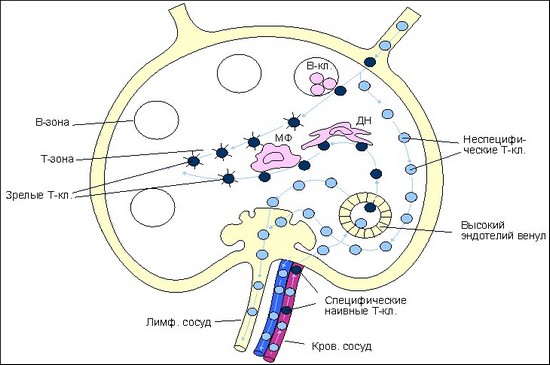
Рециркуляция Т-клеток после проникновения антигена в лимфоидную ткань.
На рисунке представлена схема лимфатического узла с направлениями движения Т-клеток по органу. Антиген с током лимфы от места проникновения в организм заносится в лимфатический узел. Антигенпрезентирующие клетки (макрофаги - МФ, дендритные клетки - ДК, В-клетки - В-кл.) захватывают, перерабатывают и представляют антигенные пептиды в комплексе с молекулами I или II классов МНС на своей поверхности. Эти события активируют процесс рециркуляции клеток.
Рециркулирующие наивные Т-клетки, среди которых имеются и специфичные к комплексу лимфоциты (черные кружки) попадают в лимфатический узел либо по афферентным лимфатическим сосудам, либо по кровеносным сосудам. В тех случаях, когда антиген поступает в орган с кровотоком, ему приходится преодолевать высокий эндотелий венул. Оказавшись в паренхиме органа, наивные антигенспецифические Т-клетки после распознавания имуногена на одной из антигенпрезентирующих клеток остаются в органе с тем, чтобы пройти дополнительную дифференцировку до функционально зрелых, армированных CD8 и CD4 Т-клеток.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Антигеннеспецифические клетки, оказавшись невотребованными, покидают орган по эфферентному лимфатическому сосуду и вступают, таким образом, в новый цикл рециркуляции.
Оказавшийся в лимфоидной ткани антиген провоцирует усиление рециркуляции лимфоцитов. Наивные Т-клетки попадают в лимфатические узлы в так называемую Т-зону через высокий эндотелий венул. Генерация зрелых (армированных) эффекторов Т-клеточного иммунного ответа начинается с распознавания антигенного пептида, комплексированного с молекулами I или II классов МНС, на поверхности макрофагов и дедритных клеток.
Сам факт распознавания комплекса является обязательным, но недостаточным условием для инициации развития наивных Т-клеток в зрелые эффекторы. Необходимо предупреждающее включение кофакторов, которые способствуют взаимодействию антигенраспознающего рецептора Т-клеток с антигенным комплексом. Именно антигенпрезентирующие клетки обеспечивают такое двойное взаимодействие.
Активация наивных Т-клеток при первичной встрече с антигеном получила название "примирование". Это определение введено с тем, чтобы отличить первичное взаимодействие с антигеном от взаимодействия зрелых эффекторов с тем же антигеном, когда собственно и проявляется функциональное предназначение различных субпопуляций Т-клеток.
В ряде случаев, в частности при формировании специфической цитотоксической реакции, презентирующая антиген клетка может выступать и как объект распознавания, и как объект цитолитического действия CD8 T-клеток после их созревания. Среди наивных Т-лимфоцитов, проникающих в лимфоидную ткань и временно локализующихся в Т-зонах, только один из 105 оказывается способным к специфическому взаимодействию. Остальные постепенно перемещаются в медуллярную зону и через эфферентный лимфатический сосуд покидают орган, чтобы вновь оказаться в кровотоке и быть занесенным в другие лимфоидные образования организма.