








Основной функцией белков главного комплекса гистосовместимости является представление антигенных пептидов в такой форме, в которой их могут распознать рецепторы Т-клеток. Как именно антигенный пептиды попадают в полость, образованную вариабельным доменами МЧС? Можно предположить, что молекулы МЧС, экспрессированных на поверхности клетки, захватывают антиген из внеклеточной среды. Однако оказалось, что молекулы МЧС без антигенного пептида в основном очень нестабильны. В 1981 г. Э. Унанью впервые убедительно доказал, что процесс связывания антигенных пептидов с МЧС происходит при активном участии клетки и требует времени и энергии. Как оказалось, ассоциация МЧС с антигенным пептидом происходит внутри клетки, и этот процесс тесно связан с биосинтеза и экспрессией молекул MHC антигенный пептиды, которые связываются с MHС образуются в результате процессинга белковых антигенов.
Процессинг - ограниченное расщепления белковых антигенов с помощью специфических протеиназ, которое происходит в клетках. Образование комплексов МЧС с антигенным пептидами и экспрессию этих комплексов на поверхности клеток называют презентации, или представлением антигенов.
Синтез молекул МНС, процессинг и представление антигенов
Молекулы МНС I класса синтезируются в грЭПС, где они формируют комплексы с антигенами, являющимися эндогенно синтезированными молекулами (например, вирусными белками в инфицироваиных клетках или белками опухолевых клеток). Эти антигены, находящиеся в гиалоплазме, предварительно подвергаются расщеплению на короткие пептидные фрагменты (8-11аминокислот) в особом протеолитическомАТФ-зависимом крупном белковом комплексе - протеасоме. Затем они транспортируются в просвет грЭПС с помощью специальных переносчиков в мембране -TAP-белков (от англ. Transporter for Antigen Presentation - переносчик для представления антигена), где связываются с молекулами МНС I класса. Образованные комплексы транспортируется через комплекс Гольджи к плазмолемме и экспрессируются на ее поверхности.
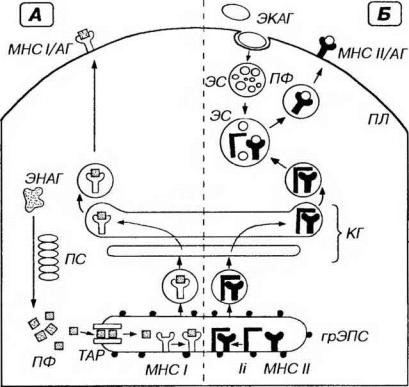
Пути процессинга и представления антигенов.
Антигены (АГ) - эндогенные (ЭНАГ) или экзогенные (ЭКАГ) - в цитоплазме АПК подвергаются процессингу, связываются с молекулами главного комплекса гистосовместимости (МНС) и экспрессируются на плазмолемме (ПЛ) в виде комплекса МНС/АГ. А. ЭНАГ в гиалоплазме расщепляются в протеасоме (ПС) на пептидные фрагменты (ПФ), которые с помощьюTAP-белков(ТАР) переносятся в просвет грЭПС. Здесь они связываются с молекулами МНС I класса (МНС I), образуя комплексы МНС 1/АГ, транспортируемые через комплекс Гольджи (КГ) к ПЛ. Б. ЭКАГ захватываются механизмом эндоцитоза и подвергаются процессингу в эндосомах (ЭС) или лизосомах до ПФ. В грЭПС молекулы МНС II класса (МНС II) и инвариантные цепи (li) связываются с образованием комплекса МНС II/ li, который через КГ направляется к ЭС, содержащим ПФ. Комплекс МНС ll/li в ЭС диссоциирует, освобождая МНС II, связывающиеся с ПФ в комплекс МНС ll/АГ, который переносится к ПЛ.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Распределение молекул МНС I класса. Молекулы МНС I класса обнаруживаются на поверхности всех клеток и тромбоцитов (благодаря чему распознающие их цитотоксические лимфоциты имеют возможность уничтожения любых зараженных или опухолевых клеток).
Молекулы МНС II класса также образуются в грЭПС, где они формируют комплекс с так называемой инвариантной пептидной цепью (Ii). Предполагают, что она препятствует связыванию молекул МНС II класса с эндогенными пептидами, является переносчиком молекул МНС II класса и содержит кодирующие сигналы для последующего направления образованного комплекса в эндосому.
Комплекс молекулы МНС II класса/цепь Ii через сеть транс-Гольджи в транспортных пузырьках направляется к эндосоме, содержащей экзогенные (например, бактериальные) антигены, предварительно подвергнутые процессингу. Пузырьки сливаются с эндосомой, внутри которой цепь Ii отсоединяется от молекул МНС II класса. Последние тут же образуют комплекс с антигенными пептидами, далее транспортируемый на поверхность клетки.
Распределение молекул МНС II класса. Гликопротеины МНС II класса экспрессируются на "профессиональных" АПК (макрофагах, дендритных АПК и В-лимфоцитах), что обеспечивает их взаимодействие с Т-хелперами. Изредка их экспрессия может индуцироваться на клетках других типов ("непрофессиональных" АПК).
Способность молекул МНС образовывать комплексы с антигенными пептидами различается у отдельных людей, что может оказывать влияние на особенности их иммунных реакций, в частности, на устойчивость к инфекциям.
Дендритные АПК, захватившие антиген, мигрируют из тканей в лимфатические капилляры, а оттуда – в Т-зависимые зоны регионарных лимфатических узлов, где они окончательно созревают и приобретают способность к представлению антигенов (в комплексе с молекулами МНС) лимфоцитам.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
При встрече с Т-лимфоцитом, обладающим рецепторами к соответствующему антигену, дендритная АПК контактно взаимодействует с ним, активируя его и инициируя развитие иммунной реакции. Характер этой реакции зависит от природы молекул МНС, связанных с антигеном. Антигены, образующие комплекс с молекулами МНС I класса, распознаются лимфоцитами с поверхностными маркерами CD8, а антигены, связанные с белками МНС II класса - лимфоцитами с фенотипом CD4 Особенности архитектоники лимфоидной ткани и путей циркуляции лимфоцитов обеспечивают максимальное количество контактов лимфоцитов с потенциальным антигеном на поверхности АПК. Более того, установлено, что дендритные АПК вырабатывают хемокин, привлекающий Т-лимфоциты.
Полноценная функция дендритных АПК способствует эффективному и своевременному распознаванию микробных и опухолевых антигенов, что препятствует развитию инфекций и новообразований. Последние часто протекают на фоне сниженной активности АПК, поэтому стимуляция функции АПК рассматривается как перспективный метод иммунотерапии таких заболеваний.